
Lifestyle Modification: The Most Powerful Treatment You Were Never Taught
Part Four: How Sleep Deprivation Dismantles Your Metabolic Health

You’ve been told to get more sleep. Perhaps you are thinking of using a pill or supplement to address your sleep problems. But, were you ever told what sleep actually does, the biological cost of poor sleep, or how you can improve sleep by adopting some specific habits?
If you haven’t, then this article is for you.

What Normal Sleep Actually Does
The American Heart Association and AASM both recommend 7 to 9 hours of sleep per night for adults (AHA, 2022). The American Academy of Sleep Medicine classifies sleep as four distinct stages cycling four to six times per night in approximately 90-minute ultradian cycles (AASM, ICSD-3-TR, 2023).
N1 is the transition from wakefulness, lasting one to five minutes. It has no significant restorative function. It is the entry stage.
N2, consolidated light sleep, is where the heart rate slows and core body temperature begins its descent. Sleep spindles appear here: bursts of oscillatory neural activity responsible for transferring memories from the hippocampus to the neocortex for long-term storage. N2 constitutes approximately 45 to 55 percent of total sleep time and is the most abundant stage in the adult sleep architecture.
N3, slow-wave sleep, is the most metabolically critical stage. The majority of nightly growth hormone secretion is concentrated in the first slow-wave episode of the night. Brain glucose utilization drops substantially. Tissue repair, immune cytokine release, and cellular restoration are concentrated here. Glymphatic clearance, the brain’s waste removal system, is maximally active during slow-wave sleep. Slow-wave sleep constitutes approximately 15 to 20 percent of total sleep time in young adults and declines substantially with age.
REM sleep produces a brain electrical pattern nearly indistinguishable from wakefulness, with skeletal muscle atonia preventing the body from acting out the activity of a near-waking brain. Brain glucose utilization returns to near-waking levels. REM is when the brain processes emotional experience, consolidates emotional memory, and performs synaptic homeostasis. The autonomic variability that occurs during REM is part of normal cardiovascular conditioning. REM episodes lengthen progressively toward morning. The final episode of a full night can last 45 to 60 minutes.
Slow-wave sleep dominates the first half, REM dominates the second. Cutting the night short at the back end disproportionately eliminates REM, while fragmenting the first half disproportionately eliminates slow-wave sleep and its attendant growth hormone secretion and glymphatic activity.
Five Factors That Disrupt Sleep Quality
Psychological Stress
Work pressure, family conflict, and occupational demand activate the hypothalamic-pituitary-adrenal axis, producing cortisol elevation and central nervous system hyperarousal that is directly incompatible with sleep initiation and maintenance. Slow-wave sleep normally exerts an inhibitory influence on the HPA axis, damping cortisol secretion. When sleep is disrupted, that inhibitory brake is removed: cortisol rises further, hyperarousal deepens, and the cycle propagates independently of the original stressor. Acute psychosocial stress increases sleep onset latency and reduces early slow-wave activity. Chronic stress sustains HPA hyperactivation associated with metabolic dysfunction, obesity risk, and cognitive impairment (Vgontzas et al., PMC2902103; PLOS Mental Health 2025 review; American Journal of Medicine 2025, doi:10.1016/j.amjmed.2025.05.044).
Irregular Sleep Scheduling
The suprachiasmatic nucleus, the brain’s master circadian clock, requires consistent sleep-wake timing to maintain biological alignment. Irregular scheduling produces social jet lag: chronic misalignment between the biological clock and socially imposed schedules. Bouman and colleagues, in a 2023 meta-analysis in the Journal of Sleep Research, found social jet lag significantly associated with metabolic syndrome parameters and type 2 diabetes risk across multiple cohort studies (Bouman et al. J Sleep Res. 2023;32:e13770). A human crossover trial demonstrated that circadian misalignment alone, independent of caloric intake and physical activity, reduced insulin sensitivity by 32% (Sládek et al. Sleep. 2023. PMC10262187). A 2024 prospective cohort study by Windred and colleagues in Sleep found sleep regularity to be a stronger predictor of mortality risk than total sleep duration, a finding with substantial implications for the widespread practice of sleeping inconsistently through the week and attempting to recover on weekends (Windred et al. Sleep. 2024;47:zsad253).
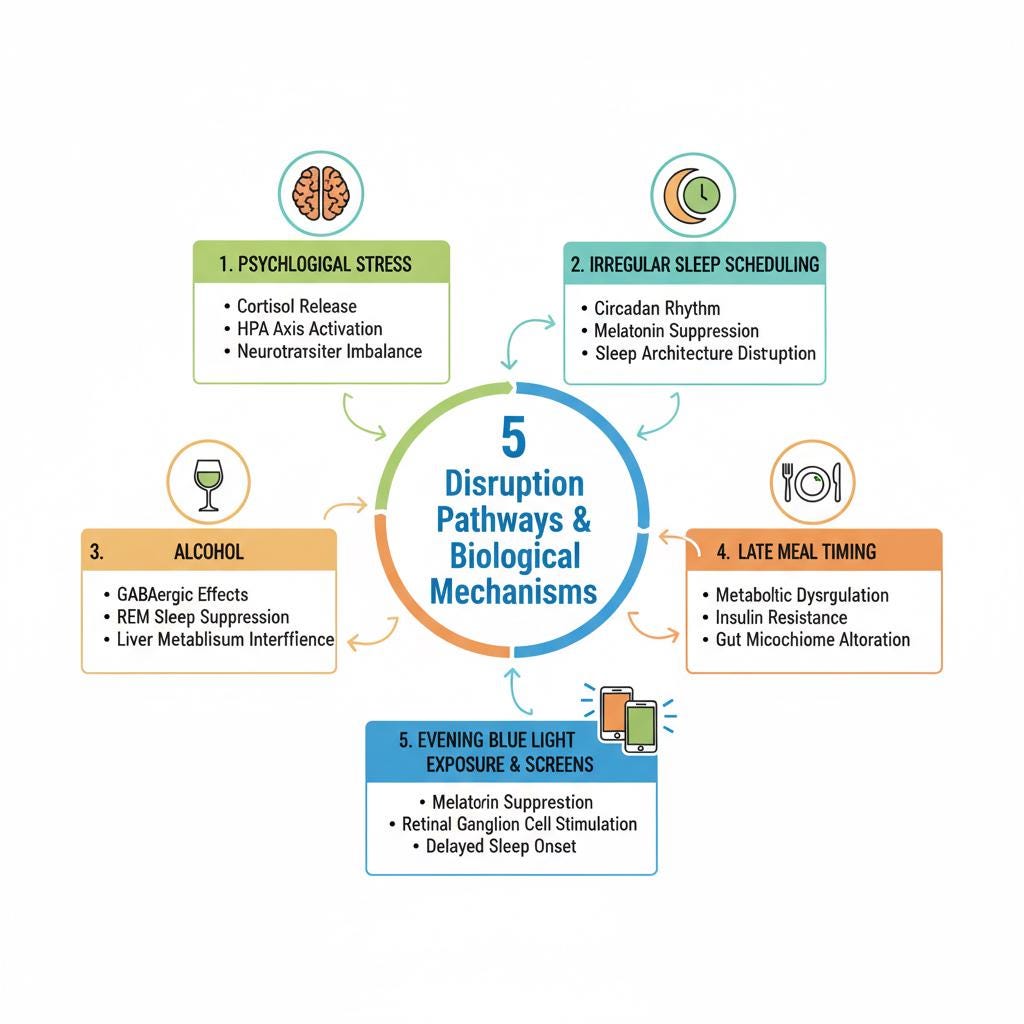
Alcohol
Alcohol is widely used as a sleep aid: alcohol shortens sleep onset latency through its effects on GABA receptors, glutamate, and adenosine pathways. The trade-off is severe and operates directly on the architecture described above. In the first half of the night, alcohol increases slow-wave sleep beyond its normal proportion while simultaneously suppressing REM. As alcohol is metabolized in the second half, its sedative effect reverses. Rebound sympathetic activation produces increased arousals, fragmented sleep, and disproportionate elimination of second-half REM, the portion of the night where REM is already most abundant and most consequential.
A 2025 systematic review and meta-analysis of 27 studies by Gardiner and colleagues confirmed a dose-response relationship: REM disruption occurs following even low doses, defined as two standard drinks or fewer, and worsens with increasing intake (Gardiner et al. Sleep Med Rev. 2025;80:102030). McCullar and colleagues demonstrated in Sleep (2024) that consecutive nights of pre-sleep alcohol produce cumulative architectural disruption that does not recover night-to-night (McCullar et al. Sleep. 2024;47:zsae003). Alcohol produces sedation while systematically dismantling the architecture that makes sleep restorative.
Late Meal Timing
Three mechanisms converge when the last meal is taken late in the evening.
First, postprandial thermogenesis generates heat that works against the core body temperature drop required for sleep initiation and slow-wave sleep maintenance.
Second, insulin sensitivity follows a circadian rhythm, peaking in the morning and declining through the evening: eating late activates insulin secretion at a point of reduced peripheral sensitivity, producing larger glycemic excursions and greater insulin demand than the same meal consumed earlier.
Third, melatonin secretion begins in the evening and suppresses insulin release from pancreatic beta cells via MT1 and MT2 receptors. A large late meal taken close to melatonin onset creates direct competition between a rising melatonin suppressing insulin and a postprandial glucose load demanding it.
A 2024 NHANES-based study in PLOS One by Yan and colleagues found that late initial, midpoint, and final meal timing were each independently associated with poor sleep quality (Yan et al. PLoS One. 2024;19(8):e0308172. PMC11293727). A 2024 systematic scoping review in Sleep Medicine Reviews confirmed the association between late eating patterns and disrupted sleep across multiple study designs (Saidi et al. Sleep Med Rev. 2024;76:101953).
Evening Blue Light Exposure and Screens
Blue light in the 460 to 480 nanometer range activates photosensitive retinal ganglion cells containing melanopsin. These cells signal directly to the suprachiasmatic nucleus, which in turn suppresses pineal melatonin secretion. The circadian system shows meaningful sensitivity to light levels below 30 lux, well within the range of a standard phone or tablet screen. Even brief evening screen use reduces melatonin and shifts its onset, delaying sleep initiation and shortening total sleep time.
A 2025 systematic review and meta-analysis in Frontiers in Neurology by Luna-Rangel and colleagues confirmed that evening blue light suppresses melatonin, delays circadian phase, and prolongs sleep onset latency (Luna-Rangel et al. Front Neurol. 2025. PMC12668929).
If you enjoy evidence-based medical information, subscribe to receive these articles delivered to your mailbox every week.
The Metabolic Consequences of Chronic Poor Sleep
Insulin Resistance and Glucose Dysregulation
Sleep deprivation activates the sympathetic nervous system and elevates cortisol, reducing peripheral insulin sensitivity by diminishing muscle glucose uptake while simultaneously enhancing hepatic glucose output. This metabolic cascade mirrors early type 2 diabetes: reduced glucose tolerance, impaired insulin-stimulated glucose disposal, and inadequate glucose-induced insulin secretion.
A 2022 meta-analysis of 21 randomized controlled trials by Sondrup and colleagues found significant reductions in insulin sensitivity across all included sleep restriction studies (Sondrup et al. 2022). One RCT found the acute insulin response during sleep restriction to be 30% lower than in well-rested conditions, a magnitude comparable to the insulin resistance associated with aging and gestational diabetes. Insomnia is associated with increased in HbA1c (Schipper et al. Diabetologia. 2021). Sleep extension of one to two weeks improved blood glucose control and insulin sensitivity in both healthy and diabetic participants across seven eligible RCTs (Kanaley et al. Exerc Sport Sci Rev. 2024;52(3):77-86. PMC11168896).
The metabolic impairment compounds across consecutive nights of restriction and is not fully reversed by a single recovery night. The person sleeping poorly across a working week is not returning to metabolic baseline on the weekend. Recovery is partial and slow.
Appetite Dysregulation and Weight Gain
Sleep deprivation disrupts hormonal appetite regulation through two documented mechanisms. Leptin, the satiety-signaling adipokine, declines with sleep restriction, removing the central inhibitory effects on appetite. Ghrelin rises, increasing appetite drive and food-seeking behavior.
Spiegel and Van Cauter demonstrated that four hours of sleep for two consecutive nights decreased leptin and increased ghrelin with quantifiable increase in self-reported hunger (Spiegel K, Tasali E, Penev P, Van Cauter E. Ann Intern Med. 2004). A controlled crossover study in 19 lean men confirmed that sleep restriction elevated 24-hour ghrelin profiles and increased snack calorie intake by 328 kcal, primarily from carbohydrates. Van Egmond and colleagues found in a 2023 Obesity study that the ghrelin response after acute sleep loss was more pronounced in individuals with obesity (Van Egmond et al. Obesity. 2023;31(3):635-641).
A 2025 meta-analysis of six RCTs found no statistically significant population-level changes in leptin or ghrelin following sleep deprivation in all studies, suggesting that central reward pathway effects are equally or more important than peripheral hormone shifts alone (Rosenberg et al. Obesity Reviews. 2026). This connects directly to the emotional dysregulation section below.
Cortisol Elevation and the Self-Perpetuating Cycle
Chronic poor sleep produces sustained HPA (hypothallamic-pituitary-adrenal) axis activation and elevated cortisol, particularly in the evening hours when cortisol should be at its lowest. Elevated evening cortisol directly impairs slow-wave sleep, the stage that normally exerts inhibitory feedback on the HPA axis, creating a self-sustaining cycle that propagates independently of the original stressor. The metabolic effects of sustained cortisol elevation are well-established: cortisol is catabolic for muscle, directly promotes hepatic gluconeogenesis, and contributes to both insulin resistance and visceral adiposity.
One RCT found that controlling for cortisol and testosterone statistically eliminated 50% of the insulin resistance induced by sleep deprivation, directly implicating hormonal dysregulation as a primary mechanism (Vgontzas et al. PMC2902103). What begins as a cause of poor sleep becomes, once chronic, a mechanism that perpetuates the disruption without any continued external stressor.
Cardiovascular Disease
Sleep deprivation and poor sleep quality are independently associated with hypertension, coronary heart disease, heart failure, atrial fibrillation, stroke, and cardiovascular mortality through converging biological mechanisms. Sustained sympathetic nervous system activation elevates resting heart rate and blood pressure. HPA-mediated cortisol elevation promotes systemic inflammation and endothelial dysfunction. Reduced slow-wave and REM sleep attenuates the normal nocturnal blood pressure dipping that protects vascular integrity. Even a single night of sleep deprivation is associated with increased arterial stiffness in healthy adults.
A 2025 PLOS One analysis found that both shorter and longer sleep durations increase the risk of heart failure, myocardial infarction, and hypertension, with optimal cardiovascular protection in the 7 to 9 hour range (PLOS One. July 2025. doi:10.1371/journal.pone.0326499). The pro-atherogenic mechanism includes sympathetic nervous system driven bone marrow release of monocytes and progenitor cells, elevated inflammatory cytokines, and endothelial adhesion molecule upregulation, all documented in human sleep deprivation studies (Franceschini et al. Nature Communications Biology. 2021. PMC8602722).
Glymphatic Failure and Dementia Risk
The glymphatic system is a brain-wide network of perivascular channels through which cerebrospinal fluid and interstitial fluid exchange, serving as the brain’s primary waste clearance mechanism. Identified by Nedergaard and colleagues in 2012 and named for its dependence on glial aquaporin-4 water channels, glymphatic activity is dramatically higher during sleep than during wakefulness, driven by reduced brain parenchymal resistance to fluid flow, reduced noradrenergic tone, and the slow oscillatory neural activity characteristic of slow-wave sleep. The primary waste products cleared include amyloid-beta and tau, the proteins whose accumulation defines Alzheimer’s disease pathology (Xie et al. Science. 2013;342(6156):373-377).
In animal models, acute and chronic sleep deprivation increase interstitial amyloid-beta levels and accelerate plaque deposition, with tau seeding and spread worsening with sleep loss. A 2024 randomized crossover trial in 39 humans directly demonstrated that overnight glymphatic clearance during normal sleep increased morning plasma levels of Alzheimer’s disease biomarkers compared to sleep-deprived conditions, confirming that the system actively clears amyloid-beta and tau to plasma in humans during sleep (PMC12847902).
The Framingham Heart Study data are the most sobering finding in this area. Himali, Pase, and colleagues followed 346 adults over 60 for 17 years and found that each percentage decrease in slow-wave sleep per year was associated with a 27% higher risk of incident all-cause dementia, adjusted for age, sex, smoking, medication use, and genetic risk (Himali et al. JAMA Neurol. 2023;80(12):1326-1333. doi:10.1001/jamaneurol.2023.3889). While only a prospective longitudinal association, is among the strongest evidence linking a modifiable sleep parameter to dementia risk over a clinically relevant time horizon. APOE e4 carrier status was independently associated with accelerated slow-wave sleep loss, suggesting that genetic Alzheimer’s risk operates partly through disruption of the very sleep stage responsible for clearing the pathological proteins.
The 2025 SLEEP Annual Meeting included formal discussion of emerging data suggesting glymphatic clearance may not be strictly sleep-dependent and may occur to some degree during wakefulness. This is an evolving area of scientific discussion. The preponderance of evidence across animal models, human RCTs, and longitudinal cohort data still supports slow-wave sleep as essential to glymphatic function and amyloid clearance (Corbali and Levey. Front Neurol. 2025. PMC11835678).
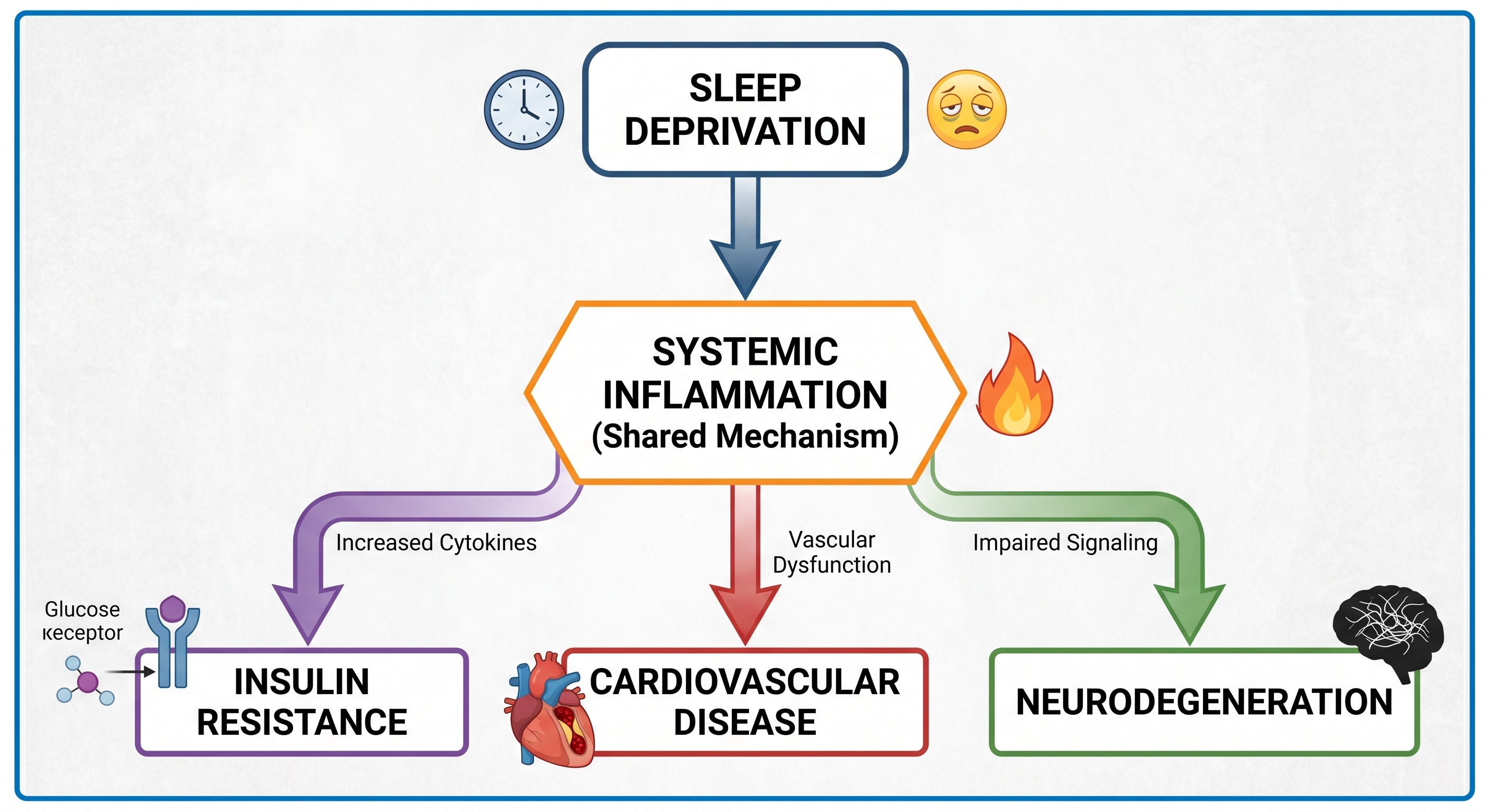
Systemic Inflammation — The Convergent Downstream Pathway
Every consequence described above shares a common amplifier: persistent low-grade systemic inflammation produced by chronic sleep deprivation.
A 2025 meta-analysis in the Journal of Sleep Research by Ballesio and colleagues, covering 35 studies and 887 participants, found that multiple nights of partial sleep deprivation, approximately 4.5 hours for three or more consecutive nights, produced significant increases in IL-6 and CRP (Ballesio et al. J Sleep Res. 2025. PMC12856123). Critically, a single night of deprivation did not produce measurable inflammatory changes. This establishes a threshold effect: the inflammatory consequence requires sustained disruption, not acute disruption. The pro-inflammatory mechanism operates through sympathetic nervous system activation increasing monocyte and neutrophil production from bone marrow, HPA-driven cortisol dysregulation altering cytokine balance, and impaired immune homeostasis across innate and adaptive compartments. Sustained inflammation is the pathway through which poor sleep accelerates insulin resistance, cardiovascular disease, and neurodegeneration simultaneously.
Acute Cognitive Impairment and Emotional Dysregulation
The prefrontal cortex is disproportionately sensitive to sleep loss. Even modest sleep restriction selectively impairs sustained attention, executive function, working memory, impulse control, and decision-making before other cognitive domains are measurably affected. A 2025 study in Frontiers in Neuroscience confirmed that sleep deprivation selectively impairs attention networks and executive function first (Ren et al. Front Neurosci. 2025. PMC12014645). A 2025 narrative review confirmed higher error rates, impaired decision-making, and reduced cognitive control across all tested domains following chronic sleep deprivation (PMC12168795).
The emotional dysregulation mechanism is the most personally recognizable consequence. REM sleep is when the brain processes and integrates emotional experience through coordinated theta-frequency oscillatory activity between the amygdala, hippocampus, and prefrontal cortex. During REM, the prefrontal cortex exerts top-down regulatory control over amygdala reactivity, allowing emotionally charged experiences to be consolidated. A 2023 Journal of Neuroscience computational modeling study confirmed that rhythmic prefrontal-amygdala interaction during REM is the basis for emotional memory processing and fear consolidation (Stickgold R, Bhatt DL, et al. Journal of Neuroscience. 2023;43(3):433-446. doi:10.1523/JNEUROSCI.1020-22.2022).
Sleep deprivation severs this regulatory circuit. Yoo and colleagues’ neuroimaging work demonstrated a prefrontal-amygdala disconnect following sleep deprivation: amygdala reactivity to emotionally provocative stimuli increased by approximately 60% while prefrontal inhibitory control over that response was simultaneously weakened (Yoo SS et al. Curr Biol. 2007;17(20):R877-8). The result is a brain operating with a hyperreactive emotional processing center and a compromised regulatory governor, producing the irritability, emotional lability, impulsivity, and heightened stress reactivity that anyone who has worked through a night shift or come off a long stretch of inadequate sleep recognizes immediately.
After a poor night, ghrelin rises and leptin falls. Simultaneously, amygdala hyperreactivity drives reward-seeking behavior while prefrontal inhibitory control is too compromised to override it. Sleep deprivation also reduces striatal dopamine D2 receptor availability, increasing the reinforcing value of food stimuli in direct parallel to the neurochemistry of substance craving (Rexrode et al. Front Sleep. 2023. PMC10624159). The person reaching for pastries at 7 a.m. after a sleepless night is not exercising poor judgment. Their brain’s regulatory architecture has been temporarily dismantled by a biological process with documented, measurable neurochemical consequences.
Now you have a better understanding of what sleep is, normal architecture and functions and the metabolic consequences of chronic poor sleep. Now that you understand the mechanisms, how can you actually improve sleep?
Part Four addresses exactly that.
Found this article useful? Share your thoughts. Join the conversation below.
Educational content on The Metabolic Archives is free, because medical information should be accessible to everyone. If you find value and want to support the work, a paid subscription is available and genuinely appreciated. Visit the About Page for additional information.
The Metabolic Archives is for educational and informational purposes only, and is not intended as medical advice, diagnosis, or treatment, and does not constitute a doctor-patient relationship. Do not adopt any recommendation discussed in any article or guides published here, make changes or abandon any prescribed medical treatment without prior consultation with your physician. Always seek the advice of your physician or other qualified health provider for any questions regarding your medical condition and recommended treatment options.
By reading this post, you acknowledge that you have read and agree to the Terms of Service of The Metabolic Archives, which govern all use of this content including restrictions on reproduction.
© 2026 The Metabolic Archives. All rights reserved.